- el momento y la frecuencia con que un determinado gen es transcripto (control transcripcional);
- el procesamiento del ARNm transcripto (control de procesamiento de ARNm);
- las moléculas de mRNA que son exportadas del núcleo al citoplasma (control de transporte del mRNA);
- los ARNm que son traducidos por los ribosomas en el citoplasma (control de traducción);
- la vida media del ARNm (control de degradación del ARNm)
- la activación e inactivación de proteínas (control de la actividad de las proteínas). De todas estas etapas de regulación, la primera es la que resulta más económica para la célula. La transcripción en los eucariotas difiere de la de los procariotas en varios aspectos.
En los procariontes, los ribosomas se unen a una molécula de ARNm en crecimiento y su traducción a una proteína comienza aun antes de que se haya completado la transcrpción. A diferencia de los anteriores la transcripción y la traducción de los eucariontes se encuentran separadas en el tiempo y en el espacio. En la transcripción de los eucariotas están implicadas tres ARN polimerasas diferentes, cada una especializada en transcribir distintos tipos de genes. Además, se requieren factores generales de transcripción, que permiten la unión de las ARN polimerasas al promotor, así como una multiplicidad de proteínas regulatorias. Además, en los eucariotas los genes estructurales no están agrupados en operones como lo están frecuentemente en los procariotas; la transcripción de cada gen se regula por separado y cada gen produce un transcripto de ARN que contiene la información codificada de un solo producto. Una vez que el núcleo se ha completado la transcripción por medio de la ARN polimerasa II, los transcriptos de ARNm (ARNm inmaduro o primario) se terminan de procesar modificando los extremos logrando las moléculas maduras antes de ser transportados al citoplasma celular a través de los poros nucleares. Este procesamiento incluye la adición
- de un casquete de metil-guanina (CAP) al extremo 5’ de la molécula cuando solo hay aproximadamente 20 pares de bases de largo. Su finalidad es proteger al ARNm de la degradación y como señal para uniese luego al ribosoma en la traducción.
- de una cola de poli-A al extremo 3’ al terminar la transcripción. Producido un clivado en un lugar específico del transcripto la poli A-polimersa , agrega una cola de adenina, generándole el extremo 3’.
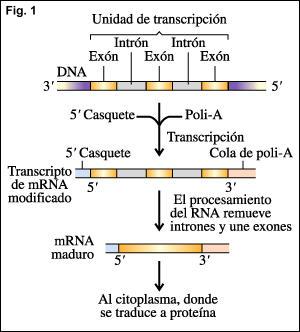
- También se produce remoción de intrones y unión de exones en el RNAm antes de dejar el núcleo. Este proceso es conocido como empalme o "splicing( Del inglés corte y empalme. ) El empalme alternativo de transcriptos de ARN idénticos en diferentes tipos de células puede producir diferentes moléculas de ARNm maduro que se traducen en diferentes polipéptidos.
Resumiendo (Fig. 1):La información genética codificada en el AND se transcribe a una copia de ARN (transcripto primario). Esta copia luego se modifica con la adición del casquete 5’ (CAP) y la cola de poli-A, la escisión de los intrones y la unión de los exones (splicing). El mRNA maduro luego va al citoplasma, donde se traduce a proteínas.Mecanismo molecular de splicing."Se trata de un mecanismo muy exacto, pues de no serlo produciría un corrimiento del marco de lectura en el mensaje transcripto. Los intrones son cortados del ARNm inmaduro por un sistema específico que reconocen secuencias cortas dentro de él y que se encuentra cerca de los límites con exones. Estas secuencias son llamadas "sitio dador" (común en casi en todos los intrones), en el extremo 5’y "sitio aceptor", en el extremo 3’ (Fig. 2). El trabajo del corte y empalme esta catalizado por una estructura pequeña, compuesta por ribonucleoproteinas nucleares llamadas snRNPs, constituidas por pequeños ARN nucleares (snARNs) asociado a proteínas. Su nombre es spliceosoma. Esta estructura tiene a su cargo el reconocimiento de las secuencias mencionadas anteriormente en los intrones y su posterior fijación. Luego se desarrollan una secuencia de pasos que determinan el clivaje y ligado de los intrones y exones (Fig. 3):
El trabajo del corte y empalme esta catalizado por una estructura pequeña, compuesta por ribonucleoproteinas nucleares llamadas snRNPs, constituidas por pequeños ARN nucleares (snARNs) asociado a proteínas. Su nombre es spliceosoma. Esta estructura tiene a su cargo el reconocimiento de las secuencias mencionadas anteriormente en los intrones y su posterior fijación. Luego se desarrollan una secuencia de pasos que determinan el clivaje y ligado de los intrones y exones (Fig. 3):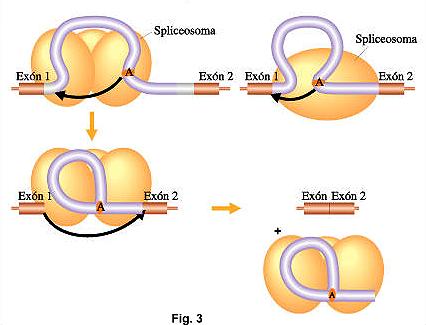
- el extremo 5’del intrón es clivado y unido a otros sitio interno del intrón, cercano a su extremo 3’ llamado "sitio de ramificación" .
- Se produce el corte en el extremo 3’ del intrón y son empalmados los dos exones de cada lado, liberándose el ARNm maduro del spliceosoma.
- El intrón eliminado queda formando una estructura con forma de lazo, llamada "lariat", que posteriormente es degradado en el núcleo.
- el extremo 5’del intrón es clivado y unido a otros sitio interno del intrón, cercano a su extremo 3’ llamado "sitio de ramificación" .
Se ha observado que ARNm inmaduros idénticos del mismo gen se procesan en más de una forma. Esto significa que existen diversos empalmes alternativos, los cuales desarrollaran diversos ARNm maduros y por lo tanto distintos polipéctidos funcionales.
No hay comentarios:
Publicar un comentario