Introducción.
La extracción de ADN requiere una serie de etapas básicas: En primer lugar tiene que romperse la pared celular y la membrana plasmática para poder acceder al núcleo de la célula. A continuación debe romperse también la membrana nuclear para dejar libre el ADN. Los jabones utilizados emulsionan los lípidos de las membranas celulares y las rompen. La sal evita la unión de las proteínas al ADN. Para aislar el ADN hay que hacer que precipite en alcohol. El ADN es soluble en agua, pero cuando se encuentra en alcohol se desenrolla y precipita en la interface entre el alcohol y el agua. Además de permitirnos ver el ADN, el alcohol separa el ADN de otros componentes celulares, los cuales son dejados en la solución acuosa.
Antecedentes.
Ademas del agua, los carbohidratos, las proteinas, y los lípidos, los seres vivos están constituidos por otras moléculas que, aunque en menor cantidad, desempeñan funciones muy importantes.
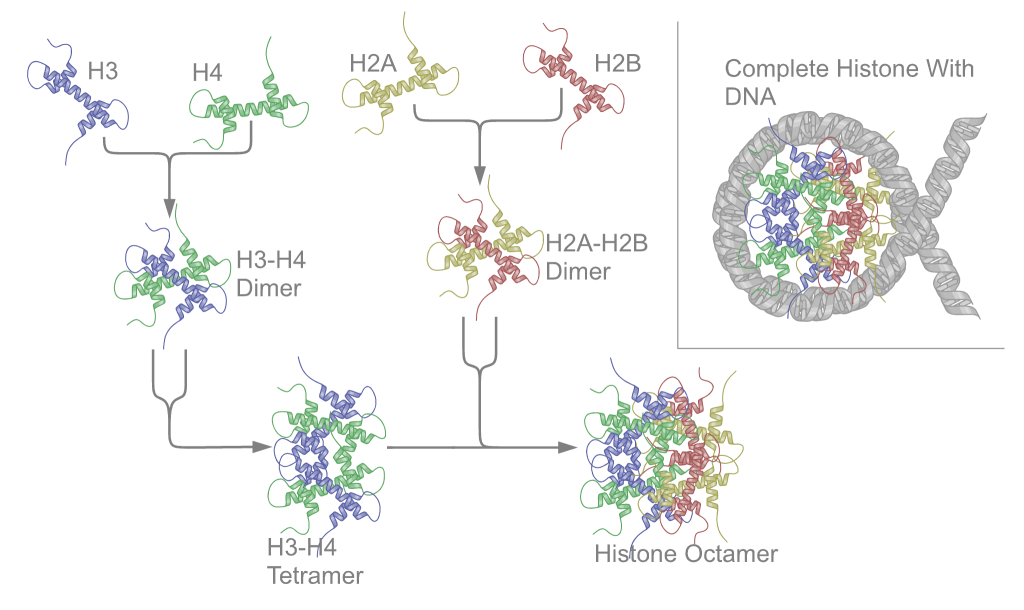
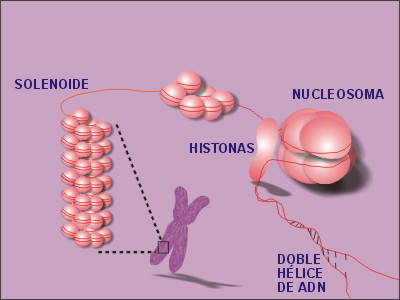
Los Ácidos nucleicos (DNA y RNA) son biomoléculas solubles en agua que desempeñan funciones diversas. Son moléculas complejas producidas por células vivas; son esenciales para todos los organismos vivos. Estas moléculas gobiernan el desarrollo del cuerpo y sus características específicas, ya que proporcionan la información hereditaria y ponen en funcionamiento la producción de proteinas.
Objetivo.
Extraer el ADN y ARN de tejidos animales o vegetales e identificarlos como tales.
Material.
Tubos de centrifuga 1ml.
Tubos de ensayo 5 ml.
Centrifuga.
Micropipetas de 1,000 y 200 ul.
Agua destilada estéril.
Etanol al 100% frio.
NaCl 3M.
Buffer de extracción.
Fenol/cloroformo/isopropanol frio.
Mortero y pistilo.
Papel aluminio.
Guantes.
Hígado de pollo.
Procedimiento.
A)._ rompimiento de membranas y paredes celulares.
Se tomo una muestra de hígado, posteriormente la muestra se homogenizó asta formar una textura de puré.
*El maceramiento del hígado es utilizado para el rompimiento de los tejidos y paredes celulares.
B)._ digestión.
En cuatro tubos para PCR se le coloco 1 gramo del hígado macerado. Para poder colocar la muestra asta el fondo se tuvo que agitar con mucha fuerza el tubo. Por ultimo se agrego 500 ul del buffer de extracción en cada tubo.
* La solución buffer impide la unión del material genético con las proteínas.
C)._ separación de proteínas, carbohidratos y lípidos.
En el tercer paso se coloco a la muestra del tubo 500 ul de fenol, con ayuda de guantes de latex y una micropipeta.
Por ultimo los cuatro tubos se colocaron en una centrifuga a 1000 rpm por un lapso de 5 min.
*la utilización de la centrifuga es para acelerar la separación de las biomoléculas. Al fondo siempre se precipitan las moléculas de mayor densidad.
D)._ precipitación de los ácidos nucleicos.
Ya terminado los 5 minutos de centrifugación, a cada uno de los cuatro tubos se le extrae la fase más superior de la muestra y se coloca en un solo tubo para PCR.
Ya extraída la fase acuosa se le coloco 400 ul de etanol y 75 ul de NaCl 3M. Pasado esto se agita vigorosamente en el Vortex.
Por ultimo se coloco el tubo en la ultracentrífuga a 1000 rpm en un lapso de tiempo de 2 min.
Conclusiones.
El proceso para la separación del material genético es un método muy meticuloso, en el cual se requiere mucha asepsia y un buen manejo de los materiales. Por el cual requiere una serie de etapas básicas.
Cuestionario.
1._ ¿Porque el ADN en solución se precipita en presencia de un alcohol frió?
R= Por que el ADN es soluble en agua, pero cuando se encuentra en alcohol se desenrolla y precipita. Ya que es insoluble en el alcohol.
2._ ¿Cómo podrías cuantificar la cantidad de DNA de tu muestra?
R= por espectrofotometría. Debidos a que los ácidos nucleicos absorben eficientemente luz ultravioleta.
3._ ¿Cuál es la función y la importancia del DNA en los organismos?
R= Estas moléculas gobiernan el desarrollo del cuerpo y sus características específicas, ya que proporcionan la información hereditaria y ponen en funcionamiento la producción de proteinas.
4._ Como se separamos de nuestra muestra el ADN del ARN?R= se realiza añadiendo enzimas que fragmentan las moléculas de ARN y que impiden que se unan al ADN.
5._ ¿Cuál es la función del cloroformo?
R= es utilizado para extraer nonatos celulares, ósea todos los componentes celulares.






















 Para separar al ADN separado con base en densidad, EL ADN se mezcla con CsCl y es centrifugado a muy alta velocidad (p. ej., 50,000 rpm) en una ultracentrifuga por muchas horas. Como los tubos giran, las fuerzas opuestas de sedimentación y difusión producen un gradiente lineal estable de CsCl con la más baja densidad arriba y la densidad más pesada en el fondo. Al formarse el gradiente de CsCl, el ADN se equilibra en el gradiente donde su densidad iguala la densidad del circundante CsCl. Si el ADN de una densidad está presente, el resultado será una sola banda de ADN. Si dos ADN están presentes con densidades diferentes, el resultado será dos bandas de ADN. Haga Click sobre el botón de "Animación" para ver una simulación de la formación de un gradiente de CsCl y la formación de una banda de ADN. Como puede ver en la la animación, note el gradiente de CsCl formado (cambio en el sombreado gris) y la formación de una banda de ADN.
Para separar al ADN separado con base en densidad, EL ADN se mezcla con CsCl y es centrifugado a muy alta velocidad (p. ej., 50,000 rpm) en una ultracentrifuga por muchas horas. Como los tubos giran, las fuerzas opuestas de sedimentación y difusión producen un gradiente lineal estable de CsCl con la más baja densidad arriba y la densidad más pesada en el fondo. Al formarse el gradiente de CsCl, el ADN se equilibra en el gradiente donde su densidad iguala la densidad del circundante CsCl. Si el ADN de una densidad está presente, el resultado será una sola banda de ADN. Si dos ADN están presentes con densidades diferentes, el resultado será dos bandas de ADN. Haga Click sobre el botón de "Animación" para ver una simulación de la formación de un gradiente de CsCl y la formación de una banda de ADN. Como puede ver en la la animación, note el gradiente de CsCl formado (cambio en el sombreado gris) y la formación de una banda de ADN.